Colaboración especial de Joan Cucurull Benet (Becario Sicómoro en el Máster del IFISC 2024-2025).
1. INTRODUCCIÓN Y CONTEXTO
El estudio de la vida (la biología) es uno de los principales focos de interés científico, entre otras cosas porque nos podría ayudar a los seres humanos a responder la pregunta de cuál es nuestra razón de ser. La biología está estructurada en diversos niveles según la complejidad del objeto de estudio, desde el nivel atómico y molecular (el más simple, dentro de la rama de la biología) hasta el nivel de ecosistema (el más complejo) pasando por el estudio de las células, los órganos, los organismos vivos pluricelulares, etc. La complejidad aumenta en cada nivel que ascendemos, ya que cada nuevo nivel está formado por objetos de los niveles anteriores y es resultado de sus interacciones. Los sistemas biológicos son claramente sistemas complejos. Cada nivel emerge de los anteriores y añade más complejidad a la estructura global de los seres vivos.
Es en este punto donde entra en juego la física de los sistemas complejos. Esta rama de la física, entre otras cosas, se dedica a investigar la interacción entre escalas de magnitud o niveles de complejidad. Por lo tanto, esta rama de la física puede ayudar a la comprensión de muchos fenómenos biológicos y al estudio en general de los sistemas biológicos. La rama que utiliza herramientas matemáticas y de la física de los sistemas complejos para el estudio de la biología se denomina biología de sistemas.
Los organismos vivos, como todo en nuestro universo tal como lo conocemos, cumplen las leyes de la termodinámica. La segunda ley, sin embargo, describe que todo tiende a un estado homogéneo y desordenado. Los seres vivos, en cambio, son el paradigma del orden frente al caos del universo. La muerte y la descomposición de los seres vivos es el paso del estado ordenado (vida) al estado desordenado al que todo tiende, implacablemente, por la termodinámica. Entender la vida es entender los mecanismos que nos mantienen vivos y que nos alejan del desorden y de la muerte.
Este proceso de mantenimiento del orden dentro de un ser vivo se produce a partir del intercambio de sustancias y de energía con el exterior. La idea es obtener energía del exterior y utilizarla para mantener el estado ordenado (la vida). Para mantener este estado hacen falta mecanismos reguladores que velen por mantener el equilibrio y la estabilidad internas: regular la concentración de sustancias, controlar las reacciones químicas, ordenar el espacio, repartir tareas, etc. Modelizar matemáticamente estos mecanismos es el principal objetivo de la biología de sistemas [2].
En este artículo exploraremos la biología de sistemas a partir de un caso paradigmático: hablaremos de algunos de los mecanismos reguladores de las células y veremos cómo se conectan la física y la biología.
2. EJEMPLO: ORGANIZACIÓN ESPACIAL, FORMACIÓN DE LA COLUMNA VERTEBRAL
Uno de los misterios de la biología es cómo las células son capaces de organizarse espacialmente y dar forma a todos los órganos y sistemas que conforman el organismo. ¿Cómo sabe cada célula cómo debe ser y dónde debe colocarse? Un ejemplo claro de organización espacial es la columna vertebral, presente en todos los animales vertebrados. Al formarse la columna vertebral, ¿cómo saben las células si pertenecen o no al tejido óseo de las vértebras, y cómo se consigue que la columna vertebral quede dividida en vértebras con una periodicidad tan perfecta?
Para responder a esta pregunta la biología de sistemas ha aportado diferentes modelos durante las últimas décadas. Estos modelos están enfocados de manera diferente según si tienen una fundamentación más química (a partir de sustancias como proteínas) o física (que se centra más en el estudio de la periodicidad espacial). En mayor o menor medida todos los modelos tienen en cuenta ambos enfoques. En la siguiente imagen vemos una cronología de los diversos modelos que han ido apareciendo en las últimas décadas:
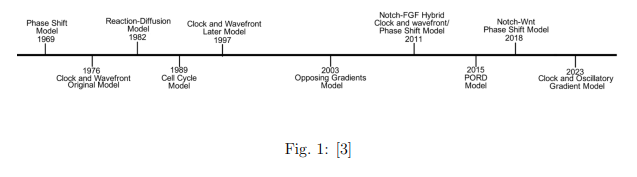
El principal objetivo de estos modelos es determinar cómo se forman las somitas: estructuras segmentadas precursoras de las vértebras de la columna vertebral. En un principio la columna vertebral es una matriz homogénea. Una parte de las células de la matriz se diferencian y pasan a formar las somitas (futuras vértebras): este es el momento crucial donde se determina la forma de la columna vertebral [3].
Los modelos mencionados en la figura anterior se basan en la idea de que la periodicidad espacial proviene en realidad de una periodicidad temporal. Esta idea aparece ya que es bien sabido que hay mecanismos por los cuales puede haber oscilaciones temporales en los seres vivos. Hay muchos ejemplos de oscilaciones en sistemas biológicos, y algunos de ellos los tenemos presentes en nuestro día a día. Un buen ejemplo son los ritmos circadianos. Nuestro estado metabólico evoluciona durante el día y este ciclo se repite periódicamente. Su presencia está regulada por factores externos como la luz del sol, pero también internamente, y es por eso que si estamos acostumbrados a levantarnos con la luz del sol a una determinada hora, si un día dejáramos la habitación a oscuras, nos levantaríamos a la misma hora ya que la regulación interna del ritmo circadiano es persistente. Siendo conscientes de la persistencia de los ciclos biológicos es lógico que las propuestas de modelos para la formación de las somitas busquen la robustez en las oscilaciones temporales: las periodicidades espaciales provienen de oscilaciones temporales biológicas.
Teniendo en cuenta la complejidad de los sistemas biológicos hay múltiples maneras por las cuales oscilaciones temporales se pueden transformar en periodicidades espaciales. Cada modelo de la figura anterior es uno diferente. En este artículo describiremos uno de los modelos más intuitivos, una versión del llamado ”Clock and wavefront model” en una versión actualizada en 2011 para encajarlo con los datos experimentales del momento [5].
El modelo se denomina ”Modelo de tres capas del reloj de segmentación”, y lo escogemos para divulgar ya que es intuitivo y nos ofrece ejemplos sobre cómo pueden interactuar diferentes niveles de complejidad, que en este caso son las tres capas.
3. MODELO DE TRES CAPAS PARA EL RELOJ DE SEGMENTACIÓN
Este modelo se llama de tres capas porque explica en tres niveles tres ingredientes imprescindibles: En primer lugar cómo puede llegar a oscilar una célula, después cómo se pueden sincronizar las oscilaciones de un conjunto de células, y para terminar cómo estas oscilaciones se trasladan a nivel espacial.
3.1. Un reloj en una célula
El principio básico de funcionamiento de las células consiste en leer el material genético y traducir su información en forma de proteínas (principales componentes funcionales de las células). Una célula puede tener un reloj interno si la concentración de alguno de sus componentes químicos presenta oscilaciones periódicas. Esto se ha visto que es posible en un circuito genético con una autorregulación negativa. Esta autorregulación aparece cuando la presencia de una proteína inhibe el proceso por el cual se crea esta proteína. Por otro lado, la proteína se va degradando por sí sola. Esto nos lleva al esquema de la siguiente figura, donde se muestra un esquema de creación de proteínas autorregulado negativamente, y un gráfico con las concentraciones de la proteína y del mRNA (la información con la cual producir la proteína).
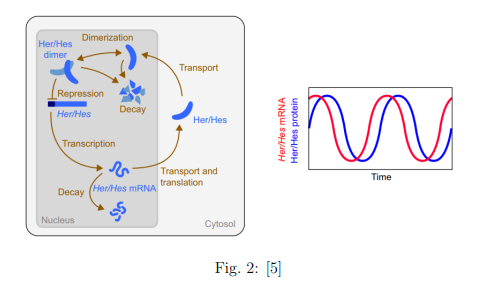
El mecanismo es claro: la producción de la proteína se reduce cuando la cantidad de proteína es alta. Por lo tanto, la cantidad de proteína acaba también reduciéndose. Esto conlleva que la producción aumente otra vez, y que la cantidad de proteína vuelva a ser alta. Este ciclo se va repitiendo periódicamente. Por lo tanto, gracias a la autorregulación negativa conseguimos que la célula se convierta en un oscilador.
3.2. Sincronización de los relojes celulares
El siguiente paso consiste en conseguir que todas las células del tejido embrionario oscilen de manera sincronizada. La manera de conseguirlo es acoplar el ciclo del apartado anterior a un ciclo más grande, también de regulación negativa, que trascienda el límite de la célula. Para realizarlo se utiliza un canal de comunicación proteico entre células que se denomina ”Delta-Notch”. Este nuevo ciclo está representado para dos células en la siguiente figura, pero es extrapolable a todo un tejido celular.
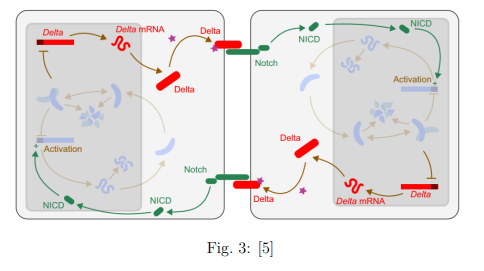
3.3. Detención de los relojes celulares
En este último paso las oscilaciones temporales se transforman en espaciales. Esto ocurre de la siguiente manera: mientras todas las células oscilan sincronizadas, pasa por ellas un frente que las deja congeladas en su estado de oscilación. Esto significa que si fueran relojes quedarían detenidos en la hora que marcaban en ese momento. Según el estado de oscilación la célula formará parte de las somitas (y por lo tanto de las futuras vértebras) o no. Según este modelo el frente viene determinado por la concentración de proteínas y factores de diferenciación celular como el FDF, el Wnt o el ácido retinoico (cuando la concentración de estas sustancias llega a un determinado umbral, los osciladores de las células se detienen y comienza la diferenciación). El frente avanza a medida que las concentraciones de estas sustancias también avanzan por el espacio. Este paso está explicado esquemáticamente en la siguiente figura:
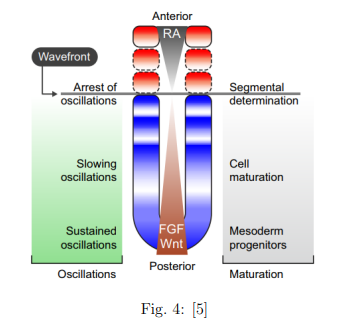
En resumen, este modelo traduce la periodicidad en el tiempo en una periodicidad en el espacio. Esto sucede de manera simple: un frente que va avanzando a través de unas células que oscilan en fase. Cuando el frente pasa a través de ellas, las células quedan congeladas en un estado concreto y dejan de oscilar. De esta manera, la oscilación temporal se ha transformado en espacial. Cabe añadir, por la evidencia experimental, que esto no ocurre de manera abrupta, sino que los relojes se ralentizan de manera paulatina y finalmente se detienen.
4. OBSERVACIÓN EXPERIMENTAL: EL PEZ CEBRA
Los tres niveles del modelo explicado anteriormente han sido observados en el proceso embrionario del pez cebra. La figura siguiente muestra una captura del proceso de formación de las somitas en un embrión de pez cebra. Podemos observar que efectivamente hay un frente que va avanzando, y que deja las somitas formadas a su paso. También describe que el período de formación de una somita es de 30 minutos (a una temperatura de 28ºC).
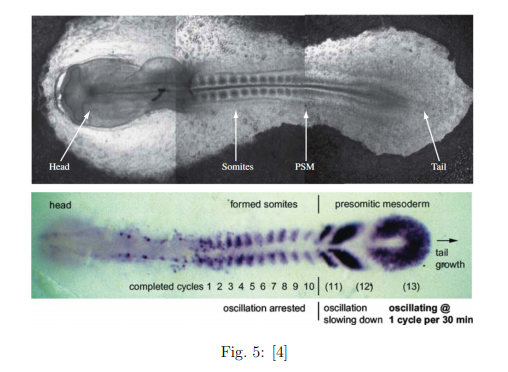
Queda bien demostrado que, tal como describe el modelo, el mecanismo de formación de patrones espaciales viene determinado por oscilaciones temporales [1].
5. CONCLUSIONES
En este artículo hemos visto cómo la física de los sistemas complejos puede allanar el camino para entender procesos biológicos de alta complejidad. La biología de sistemas nos ayuda a entender la vida con modelos matemáticos. En este caso lo hemos aplicado al proceso de formación de las vértebras en embriones. Comprender la formación de los embriones en animales vertebrados nos puede ayudar a detectar malformaciones y enfermedades de manera prematura y poder paliarlas a tiempo. No obstante, esta línea de investigación sigue abierta ya que todavía hay muchas preguntas por responder. Faltan piezas por encajar a nivel molecular y bioquímico, pero también a nivel general: ¿con qué mecanismo se controla el número total de vértebras?
Hay que seguir investigando y mejorando los modelos, pero en todo caso con el conocimiento actual podemos entender intuitivamente cuál es el mecanismo que ha ideado la naturaleza para poder conseguir unos patrones espaciales tan perfectos como son las vértebras de nuestras columnas vertebrales que nos mantienen erguidos cada día.
La biología de sistemas es una rama de la física de los sistemas complejos que todavía se encuentra en desarrollo. Según mi opinión es una de las ramas más interesantes a nivel de conocimiento, ya que estudia la vida de manera general y por lo tanto ayuda a responder preguntas filosóficas como quién somos y por qué existimos. Por otro lado, el estudio de la biología y sobre todo del cuerpo humano siempre es bienvenido dentro y fuera de la comunidad científica, ya que tiene efectos sobre nuestra vida (nos permite curar enfermedades y tener más calidad de vida). Es por eso que creo que en los próximos años seguirá creciendo la cantidad de recursos destinados a investigar en esta dirección.
¡Habrá que irlo siguiendo!
REFERENCIAS
[1] François Giudicelli et al. “Setting the Tempo in Development: An Investigation of the Zebrafish Somite Clock Mechanism”. In: PLOS Biology 5.6 (May 2007), pp. 1–15. doi: 10.1371/journal.pbio.0050150. url: https://doi.org/10.1371/journal.pbio.0050150.
[2] L. Konieczny, I. Roterman-Konieczna, and P. Spólnik. Systems Biology: Functional Strategies of Living Organisms. Springer International Publishing, 2023. isbn: 9783031315572. url: https://books.google.es/books?id=IlXEEAAAQBAJ.
[3] Cassandra Mcdaniel, Muhammed Simsek, and Ertugrul Ozbudak. “Spatiotemporal control of pattern formation during somitogenesis”. In: Science Advances 10 (Jan. 2024). doi: 10.1126/sciadv.adk8937.
[4] Philip J. Murray, Philip K. Maini, and Ruth E. Baker. “The clock and wavefront model revisited”. In: Journal of Theoretical Biology 283.1 (2011), pp. 227–238. issn: 0022-5193. doi: https://doi.org/10.1016/j.jtbi.2011.05.004. url: https://www.sciencedirect.com/science/article/pii/S0022519311002384.
[5] Andrew C. Oates, Luis G. Morelli, and Saúl Ares. “Patterning embryos with oscillations: structure, function and dynamics of the vertebrate segmentation clock”. eng. In: Development (Cambridge) 139.4 (2012), pp. 625–639. issn: 0950-1991.
